Eolambiya - Eolambia
| Eolambiya | |
|---|---|
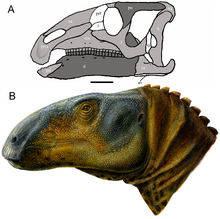 | |
| Bosh suyagini tiklash (A) va hayotni tiklash (B) | |
| Ilmiy tasnif | |
| Qirollik: | Animalia |
| Filum: | Chordata |
| Klade: | Dinozavrlar |
| Buyurtma: | †Ornithischia |
| Suborder: | †Ornitopoda |
| Superfamily: | †Hadrosauroidea |
| Klade: | †Hadrosauromorf |
| Tur: | †Eolambiya Kirkland, 1998 |
| Tur turlari | |
| †Eolambia caroljonesa Kirkland, 1998 yil | |
Eolambiya ("tong" degan ma'noni anglatadi lambozaurin ") a tur o't o'simliklari hadrosauroid dinozavr boshidan Kechki bo'r ning Qo'shma Shtatlar. Unda bitta tur mavjud, E. karoljonesatomonidan nomlangan paleontolog Jeyms Kirkland 1998 yilda turdagi namunalar ning Eolambiya 1993 yilda Kerol va Ramal Jons tomonidan topilgan; tur nomi Karolni sharaflaydi. O'shandan buyon skeletning deyarli barcha elementlarini ifodalovchi kattalar va balog'atga etmagan bolalardan yuzlab suyaklar topilgan. Hozirgacha barcha namunalar topilgan Emeri okrugi, Yuta, Mussentuchit a'zosi sifatida tanilgan tosh qatlamida Sidr tog'ining shakllanishi.
Uzunligi 6 metrgacha (20 fut), Eolambiya o'z guruhining katta a'zosi. U Osiyo hadrosauroidlariga yaqinlashganda Equijubus, Probaktrosaurus va Choyrodon, Boshsuyagi xususiyatlarida, umurtqalar va oyoq-qo'llari, aslida Shimoliy Amerika bilan chambarchas bog'liq bo'lishi mumkin Protohadros. Ning to'g'riligiga asoslangan bu guruhlash to'rtburchak suyagi va skapula, izolyatsiyani anglatadi, endemik nurlanish hadrosauroidlar. O'xshashiga qaramay hadrosauridlar - xususan, lambozozinli hadrosauridlar - bir nechta xususiyatlarga ko'ra, ularning biri sifatida dastlabki identifikatsiyasiga olib keladi, bu o'xshashliklar umuman rad etildi yaqinlashuvchi yoki noto'g'ri talqin qilingan.
Eolambiya ko'llar bo'yidagi o'rmonli muhitda namlikda yashagan bo'lar edi toshqin suv toshqini atrof-muhit, ovqatlanish gimnospermlar, ferns va gullarni o'simliklar. Vaqt o'tishi bilan ko'llardagi suv sathlari davriy nam va quruq sehrlar bilan o'zgargan oldingi Mussentuchit a'zosining cho'kindilaridagi o'zgaruvchan tasmalar bilan aks etgan Yerning Voyaga etmagan bola sifatida Eolambiya katta o'lja edi krokodilomorflar ko'l suvlarida yashovchi. Yoshi o'sib borishi bilan ular krokodilomorflarga beparvo bo'lib qolishdi va etuk shaxslar (kamida sakkiz-to'qqiz yosh) katta o'lja bo'lishdi. tropodlar kabi neovenatorid Siats.
Kashfiyot tarixi
Dastlabki kashfiyotlar va tavsif

1979 yilda Piter Galton va Jeyms A. Jensen parchalangan o'ng femur tasvirlangan, BYU Ga tegishli bo'lgan 2000 yil hadrosaurian ga tegishli cho'kindi jinslardan topilgan dinozavr Sidr tog'ining shakllanishi yilda Arches milliy bog'i, Yuta. Kambag'al material bo'lsa-da, u uchun muhim edi (qog'ozda tasvirlangan ikkinchi Shimoliy Amerika femurasi bilan bir qatorda) bu birinchi hadrosaur namunasi edi. Pastki bo'r dunyoning istalgan nuqtasida. Galton va Jensen kelajakda shakllanish natijasida hadozavrning to'liq qoldiqlarini topish mumkin deb taxmin qilishdi.[1] G'arbiy mintaqadagi mayda umurtqali hayvonlar karerlarida turli xil hadrosauroid tishlar topilgan San Rafael Shvell, yaqin Dale qal'asi yilda Emeri okrugi, Yuta; ular 1991 yilda J. Maykl Parrish tomonidan tasvirlangan. Keyinchalik, 1993 yilda Kerol Djons va uning eri Ramal Jons shishganlikning shimoli-g'arbiy qismida joylashgan toshqotgan joydan parchalangan suyaklarni topdilar. Ular saytni keyinchalik nomlangan muassasa direktori Donald Burjning e'tiboriga havola etishdi Sharqiy Yuta prehistorik muzeyi kolleji (CEUM). Rasmiy ravishda CEUM Locational 42em366v deb nomlanuvchi sayt keyinchalik Kerol sayti (sic ) uning sharafiga. CEUM 9758 namuna raqami ostida saqlangan qoldiqlar bosh suyagi, umurtqa pog'onasi, iskiya va oyoq qismlarini o'z ichiga olgan kattalar hadrosauroidining qisman qoldiqlarini anglatadi. CEUM 5212, qisman bosh suyagi va kattalarning old qismi, yaqin atrofdagi CEUM Mahalliyidan 42em369v topilgan.[2] CEUM 8786, kattalar chap suyagi, keyinchalik Kerol saytida topilgan va 2012 yilgacha ta'riflanmagan.[3]
Shu bilan birga, Oklaxoma tabiiy tarix muzeyi (OMNH) Shishganning janubi-g'arbiy mintaqasida olib borilgan qazishmalar natijasida xuddi shu hadrosauroidga tegishli qoldiqlarni topdi.[2] OMNH tomonidan kashf etilgan namunalar dastlab beshta joydan oltitadan iborat edi: ikkita balog'atga etmagan bolalar skeleti,[4] shu jumladan OMNH v237 joyidan vertebra, skapula, ulna, ilium va ischium va orqa elementlar; OMNH v824 dan orqa miya umurtqasi bilan birga bosh suyagi, old oyoq va orqa oyoq qismlarini o'z ichiga olgan qisman balog'at yoshidagi skelet; OMNH 27749, OMNH v696 dan sakrum va iskiyum; OMNH 24389, OMNH v214 dan iskiyum; va OMNH 32812, qisman skelet, shu jumladan skapula, ikkita dumaloq vertebra va OMNH v866 dan boshqa qazib olinmagan elementlar. Shuningdek, OMNH turli xil karerlarda hadrosauroid tishlarini kashf etdi. OMNHdan Richard Cifelli ruxsat berdi Jeyms Kirkland to'plangan namunalarni o'rganish.[2] Kirkland 1994 yilda o'tkazilgan yillik yig'ilishda konferentsiya avtoreferatida oldindan namunalarni tasvirlab berdi Umurtqali hayvonlar paleontologiyasi jamiyati.[5]

Byulletenida chop etilgan 1998 yilda chop etilgan maqolada Nyu-Meksiko tabiiy tarix va fan muzeyi, Kirkland rasmiy ravishda namunalarni yangi deb nomladi tur, Eolambiya, bilan turi va faqat mavjudotlar Eolambia caroljonesa. Umumiy ism Yunoncha prefiks eos/ ἠώς ("tong", "tong", "erta" degan ma'noni anglatadi) qo'shimchasi bilan lambiya (olingan Lambeosaurus, bu esa o'z navbatida nomlangan Kanadalik paleontolog Lourens Lambe ). Umuman olganda, nasab nomi "tong otish (yoki erta)" degan ma'noni anglatadi. lambozaurin ", uning eng bazal lambeozaurin pozitsiyasiga nisbatan. Shu bilan birga aniq ism Carole Jonesni sharaflaydi. Ism Eolambiya tomonidan taklif qilingan paleoartist Maykl Skrepnik; u 1998 yilgi qog'ozdan oldin Kirkland tomonidan ishlatilgan "Eohadrosaurus caroljonesi" norasmiy nomini almashtirdi.[2]
Keyingi kashfiyotlar

1998 yildan beri olib borilgan qo'shimcha qazishmalar natijasida skeletga oid qo'shimcha elementlar aniqlandi Eolambiya. Hozir bu eng taniqli iguanodont hisoblanadi Senomiyalik va Santonian bosqichlar. Taxsonga tegishli deyarli barcha skelet elementlari mavjud kollektsiyalarda bir nechta toshqotganliklar bilan ifodalanadi. Kamida o'n ikki kishi, shu jumladan kamida sakkizta voyaga etmaganlar,[6] shahri janubidagi karerda topilgan Emeri ikkala bosh suyagi va tananing qolgan qismlarini o'z ichiga oladi. Karer Cifelli # 2 nomi bilan mashhur Eolambia caroljonesa Karer, yoki rasmiy ravishda CEUM joyi 42em432v. Emeri shahridan janubdagi yana bir karer - Willow Springs 8 karerasi yoki CEUM 42Em576v joyi - kamida to'rt nafar balog'atga etmagan bolani saqlaydi. Ushbu karerlarning materiallari 2012 yilda Kirkland tomonidan Endryu Makdonald, Jon Bird va Piter Dodson bilan birgalikda tasvirlangan, ular ham yangilangan tashxis uchun Eolambiya ushbu material asosida.[3]
Cifelli №2 karerida topilgan bitta namunadir, CEUM 34447 sifatida kataloglangan o'ng tish stantsiyasi, boshqasiga nisbatan g'ayrioddiy Eolambiya namunalar, shu jumladan kattalar va balog'atga etmagan bolalar. Boshqa odamlarda tish protezlarining o'rtacha nuqtasi va umumiy uzunligi o'rtasidagi nisbat odatda 0,19 dan 0,24 gacha bo'lsa, CEUM 34447 da xuddi shu qiymat 0,31 ga teng bo'lib, uni g'ayrioddiy darajada chuqurlashtiradi. Tish tishi, shuningdek, old tomondan sezilarli darajada chuqurlashmaydi va shu bilan ajralib turadigan xususiyatlardan biri yo'q Eolambiya tish shifokorlari. Shu bilan birga, koronoid jarayonining oldingi uchini kengaytiradigan boshqa ajralib turadigan xususiyat namunada mavjud. Garchi bu shaxs g'ayrioddiylikni anglatsa ham Eolambiya namuna, McDonald va uning hamkasblari bu farqli hadrosauroidni ko'rsatishi mumkinligi haqida ogohlantirdi morfologiya dan tish shifokorining Eolambiya (karerdagi boshqa materiallar tomonidan tavsiya etilmagan bo'lsa ham). Shunday qilib, ular buni noaniq bazal hadrosauroid deb hisoblashdi.[3]

The Tabiat tarixi dala muzeyi (FMNH) 2009 yildan 2010 yilgacha Emeri shahri janubida qazish ishlarini olib borgan Yuta geologik xizmati. Ushbu qazish ishlari Akiko Shinya tomonidan 2008 yilda topilgan, FMNH joyi UT080821-1 bo'lib, uning sharafiga Akikoning sayti deb nomlangan. Qazish ishlari paytida 167 ta disartikulyatsiya qilingan, ammo bir-biri bilan chambarchas bog'liq bo'lgan elementlar ikkita gips ko'ylagi ichida to'plangan. Kestirib, umurtqadan elementlarni o'z ichiga olgan ushbu gips kurtkalardan biriga FMNH PR 3847 raqam raqami berilgan; namuna keyinchalik McDonald tomonidan Terri Geyts, Lindsay Zanno va Piter Makoviksi bilan birgalikda 2017 yilda tasvirlangan. FMNH tomonidan kashf etilgan yana bir joy - FMNH UT130904-2 uch qavatli kareridir, unda 400 dan ortiq balog'at yoshiga etmagan suyaklarning suyagi bor. kamida to'rt kishiga.[7]
Geologiya va tanishish

Barcha joylar Eolambiya Mussentuchit a'zosiga tegishli bo'lganligi aniqlandi Sidr tog'ining shakllanishi. Mussentuchitdagi ustun jinslar bir-biridan ochilgan kulrangdan kulranggacha bo'lgan qatlamlardan iborat loyli qumtosh va loy toshi. Ikkala alohida depozitlar daryo kanali tizimlar Mussentuchit-da ham mavjud. Ushbu tizimlarning pastki qismi o'z navbatida loyli qumtosh, mayda donali qumtosh, loyqa oxirgi ikki qatlamda o'simlik qoldiqlari bo'lgan loy va mayda donali qumtosh. Cifelli №2 karer ushbu tizim bilan bog'liq. Yuqori tizim yam-yashil mayda donali qumtoshdan iborat. Uch qatlam bentonit gil vakili qiluvchi qatlamlar vulkanik kul konlari, shuningdek, oq-kulrang, sariq va sarg'ish rangda bo'lgan Mussentuchitda mavjud. Ushbu qatlamlarning o'rtasi Cifelli # 2 karerini kesib o'tadi.[6] Bentonit, shuningdek, Mussentuchit bo'ylab qumtoshning o'zida mavjud.[7] Cifelli №2 karerida suyaklar o'zlari bilan qorayib, qoraygan kaltsit, kvarts, pirit va karbonat -gidroksiapatit.[6]
Palinologiya (polenni o'rganish),[8][9][10] shuningdek, fotoalbomlarni o'rganish megaflora va umurtqasizlar,[11] dastlab Sidr tog 'shakllanishining eng yuqori qismida, shu jumladan Musentuchitda, - deb taxmin qilgan edi Albian bosqich.[2] 1997 yil argon-argon bilan tanishish ning sanidin Cifelli va uning hamkasblari tomonidan yuqori Sidar tog'lari shakllanishida vulkanik kul qatlamidan tiklangan kristallar 97 ± 0,1 million yilni ko'rsatdi, bu keyinchalik Albian va Senomanian bosqichlari chegarasidan keyin deb hisoblanadi. Bu ortiqcha senomian yoshiga o'xshaydi Dakota shakllanishi.[12] Keyinchalik, 2007 yilda Jeyms Garrison va uning hamkasblari Cifelli №2 kareridan o'tuvchi vulkanik kul qatlamini belgilashdi - bu ehtimol Cifelli va uning hamkasblari tomonidan ilgari tuzilgan qatlam - 96,7 ± 0,5 million yil oldin. Taxminan darajada Cifelli va Garrison taxminlari bir-biriga mos keladi, shuning uchun senomaniyalik yoshni bildiradi Eolambiya. Garrison va uning hamkasblari, shuningdek, Mussentuchit bo'ylab joylashgan joylardan polen to'planishini qayta tahlil qilib, yig'ilish Albian-Cenomanian chegaralarini qamrab olgan degan xulosaga kelishdi.[6]
Tavsif

Eolambiya katta hadrosauroid. Dastlabki hisob-kitoblarga ko'ra uning bosh suyagi uzunligi 1 metrga (3 fut 3 dyuym) teng bo'lgan,[2] garchi bu nomutanosib uzun burun tufayli bo'lsa-da, keyinchalik qo'shimcha materiallar topilishi bilan tuzatilgan.[3] 2016 yilda, Gregori S. Pol tana uzunligi 6 metr (20 fut) va og'irligi 1 metrik tonna (0,98 uzun tonna; 1,1 qisqa tonna) uchun taxmin qilingan Eolambiya,[13] tanasining uzunligi 6,1 metr (20 fut) bo'lgan oldingi taxminiga muvofiq keladi Tomas R. Xolts Jr. 2012 yilda.[14] Avvalroq, 2008 yilda kattalar namunasi uzunligi 5,2 metr (17 fut) va balandligi 2 metr (6 fut 7 dyuym) balandlikda deb taxmin qilingan edi.[6]
Boshsuyagi

Bezsiz bosh suyagi Eolambiya ga o'xshash umumiy shaklga ega Equijubus va Probaktrosaurus. Tuproqning old qismi juda qo'pol bo'lib, ko'pchilik tomonidan punktuatsiya qilingan foramina (teshiklar). Har birining uchida premaxilla, dentikulalar deb nomlanuvchi ikkita tishga o'xshash tuzilmalar mavjud bo'lib, ular ham eng yaqin qarindoshida ko'rinadi Protohadros. Oldinga orqada, preaksillaning pastki qismining orqa qismi to'satdan yuqoriga ko'tarilib, orqadagi burun teshigini yopilgandek yopadi. Probaktrosaurus, Protohadros, va boshqa hadrosauroidlar. Ushbu qism ikkita barmoqqa o'xshash jarayonlarga qo'shiladi maxilla, shunga o'xshash Protohadros. Maksillaning tanasi chuqurchaga ega emas yoki har qanday ko'rsatgichga ega emas antorbital fenestra, kabi Equijubus, Protohadros, va boshqa hadrosauroidlar. Ajratish uchun ishlatiladigan xususiyatlardan biri Eolambiya yon tomondan qaralganda maksillaning tish qatoridagi konkav profilidir, shunga o'xshashdir Equijubus, Probaktrosaurusva boshqa bir nechta narsalar hadrosauriformalar ammo farqli o'laroq Protohadros.[3]
Yoqdi Probaktrosaurus va boshqa hadrosauroidlar, maxilla orqa tomoni bilan bog'lanadi jugal - bu ko'z bo'shlig'ining pastki qismi bilan chegaralanadi va zamonaviy bo'lmagan fenestra - chuqurchaga to'g'ri keladigan barmoqqa o'xshash proektsiya orqali. Jugalning pastki chekkasida infratemporal fenestra sathi ostida kuchli gardish bor; bu ham ko'rinadi Equijubus, Probaktrosaurus, Protohadros, va boshqa bir qancha hadrosauroidlar. Jugalga yuqoridan ulanish bu postorbital, bu ko'zning teshiklari bilan chegaralangan qo'pol sirtga ega (shunga o'xshash) Protohadros), ammo suyakning yon tomoni aks holda silliqdir. Bosh suyagining orqa qismida kvadrat bilan ifodalaydi skuamozal yuqoridan qaralganda D shaklidagi bo'g'in bilan. Chap va o'ng skuamosallar bir-biri bilan juda ko'p aloqa qilishgan bo'lar edi, faqat orqada faqat kichik jarayon parietal. The supraoksipital suyak Boshsuyagi orqa qismining yuqori qismini tashkil etuvchi, xuddi shunday holatdagi kabi, tekis va deyarli vertikaldir Probaktrosaurus va boshqa hadrosauroidlar.[3]

Premaxilla bilan bo'lgani kabi oldingi ning Eolambiya tish teshiklari. Bu erda taniqli dorsomedial jarayon mavjud, shuningdek yorliqqa o'xshash tuzilish Probaktrosaurus va boshqa hadrosauriformalar. Dorsomedial jarayonning ikkala tomonida bir nechta qo'shimcha yorliqqa o'xshash dentikullar mavjud edi, ular xuddi shunday Probaktrosaurus. Oldingi orqa tomonda stomatologik pastki jag'ning ko'p qismini tashkil qiladi. Qisqa tanaffus mavjud, yoki diastema, dentary bilan tishli tish bilan birinchi tish holati o'rtasida artikulyatsiya o'rtasida kuzatiladi. Equijubus, Probaktrosaurus, va boshqa hadrosauroidlar. Tish tishining old qismi xarakterli ravishda chuqurlashadi, xuddi Protohadros, Ouranosaurus va Bactrosaurus. Tish tishining tashqi yuzasida ikkita bo'rtma mavjud bo'lib, ulardan biri koronoid jarayon kabi Probaktrosaurus va boshqa hadrosauroidlar. Yoqdi Protohadros va boshqa bir qancha hadrosauriformalar, ammo farqli o'laroq Probaktrosaurus, faqat ushbu jarayonning oldingi uchi qalinlashadi. Xuddi shunday Equijubus, Probaktrosaurusva boshqa har xil narsalar iguanodonts, yon tomonida kichik teshik mavjud burchakli dentary orqasida joylashgan.[3]
Boshqa iguanodontlarga o'xshash tishlarning Eolambiya bir-biriga chambarchas bog'langan va bir-biriga bog'langan qatorlarga joylashtirilgan. Har qanday vaqtda, 32 ta yuqori tish cho'ntagining har biri uchta tishni ushlab tursa, 30 ta tish tishli uyasining har biri to'rtta tishni ushlab turadi. Ulardan har bir rozetkaning ikkitasi tishlarga o'xshash o'rnini bosuvchi tishlardir Probaktrosaurus; boshqalari faol tishlardir. Har bir faol tishning bittasi bor yuz kiymoq. Yoqdi Probaktrosaurus, Gongpoquansaurus, ProtohadrosVa boshqa hadrosauroidlar, har bir tish tish tojida faqat bitta tizma bor, u og'izning o'rta chizig'iga ozgina siljiydi. Ayni paytda, har bir tish shifokori tish toji xarakterli ravishda boshlang'ich tizma va og'izning o'rta chizig'iga yaqinroq bo'lgan aksessuar tizmasi mavjud, bu holat ham mavjud Protohadros va boshqa hadrosauroidlar. Shuningdek, barcha tojlarda old va orqa qirralarda dentikula o'xshash kichkina serralar bor, bu ham Probaktrosaurus.[3]
Umurtqa pog'onasi

Yoqdi Equijubus, Probaktrosaurus, Gongpoquansaurusva boshqalar styracosternans, Eolambiya bor bachadon bo'yni umurtqalari juda opisthocoelous, ya'ni oldingi uchlari kuchli konveks, orqa uchlari esa kuchli konkav degan ma'noni anglatadi. G'ayritabiiy tarzda, uchinchi bo'yin bachadonining orqa uchi orqa tomondan biroz yuqoriroq qilib o'rnatiladi, bu boshqa bo'yin bachadon umurtqalarida ko'rinmaydi. Bo'yinning orqa qismidagi vertebra cho'zinchoq o'rtasida katta burchakka ega qo'shma jarayonlar postzigafofizlar deb nomlanuvchi va yana cho'zilgan ko'ndalang jarayonlar. Transvers jarayonlarda yana artikulyar jarayonlar mavjud - dumaloq parafofizlar va tayoqchaga o'xshash diafofizlar. Orqa bachadon bo'yni qismida postzigafofizlarni ajratuvchi chuqur depressiya va asab tizmalari tik burchakli oldingi chekkalarga ega, bu esa boshoqqa o'xshash taniqli proektsiyalarni yaratishga ta'sir qiladi.[3][7]

Birinchi bir nechta orqa umurtqalari bachadon bo'yni umurtqalariga o'xshash, ammo uzunroq va uzunroq o'xshash nerv nervlari. Qolganlari amfiplatyan, ya'ni ikkala uchi tekis. Bundan tashqari, ular uzun bo'yli va to'rtburchaklar nerv umurtqalaridan tashqari kamroq cho'zilgan va postamentga o'xshash postzigafofizlarga ega. Proektsiyalar o'rniga depressiyalar bo'lgan parafofizlar ko'ndalang jarayonlardan to ga o'tdilar asab kamari, ko'ndalang jarayonlar va vertebra old qismidagi prezigapofizlar deb nomlanadigan artikulyar jarayonlar o'rtasida. Biroq, ular so'nggi bir nechta dorsallarda transvers jarayonlar bazasiga qaytib boradilar.[3] Uchinchi yoki to'rtinchi dorsalda parapofiz juda yaqin joylashgan tikuv asab orqa miya bilan tsentrum, bu boshqa dorsallarga o'xshamaydi, lekin undagi dastlabki dorsallarga o'xshaydi Edmontosaurus. Shuningdek, shunga o'xshash Edmontosaurus, prezigafofizlarni ajratib turadigan median tizma orqa dorsallarda yanada aniqroq bo'ladi.[7]
Orasida Eolambiya hozirgi kungacha topilgan namunalar, eng yaxshi saqlanib qolgan sakrum etti umurtqani o'z ichiga oladi. Ushbu shaxsning etuk emasligi va hadrosaurlarning yoshi bilan sakral vertebra sonini ko'payishini hisobga olsak, kattalar ko'proq sakralga ega bo'lishlari mumkin.[2] Kaudal (quyruq) umurtqalari amfikoel, old va orqa uchlari konkavga ega. Ularda postamentga o'xshash prezigapofizlar va tabga o'xshash postzigapofizlar mavjud bo'lib, ularning ikkinchisi depressiya bilan ajralib turadi. Prezigapofizlar cho'zilib, poyaga aylanadi va postzigafofizlar o'rta va orqa kaudallarda qisqaradi. Bundan tashqari, asab nayzalari to'rtburchaklar shaklidan kuchli kavisli tomonga o'tadi (old tomonida konkav va orqada konveks), markazlar yanada cho'zilib ketadi va ko'ndalang jarayonlar ham yo'qoladi.[3][7]
Oyoq-qo'llar va oyoq-qo'llar kamarlari

Yoqdi Probaktrosaurus, skapular pichoq ning Eolambiya deyarli to'rtburchaklar shaklida, pichoqning yon tomonlari to'g'ri va pichoqning uchini deyarli o'ng burchak ostida uchratadi. The humerus o'rta chiziqqa egilib, humerusdagi deltopektoral tepalik sezilarli darajada qalinlashgan. Bilan bog'lanishning tashqi yuzasida tugma mavjud radius humerusda. Ikkalasi ham ulna va radiusi nisbatan to'g'ri suyaklardir. Ulnaning yuqori qismida uchta "tish" bor, ya'ni olekranon jarayoni va ikkita qo'shimcha uchburchak gardish. Ayni paytda, radius yuqori uchida tekislangan ichki yuzani va pastki uchida ichki yuzada alohida platformani, ikkalasi ham ulna bilan artikulyatsiya qilish uchun. Qo'lda beshta raqam bor. Orasida metakarpal suyaklar qo'lning, ikkinchisi kavisli, uchinchisi to'g'ri, to'rtinchisi kuchli egri. Yoqdi Iguanodon, Probaktrosaurusva boshqalar bazal iguanodonts, birinchi raqamdagi tirnoq aniq konus shaklida bo'ladi. Ikkinchi va uchinchi tirnoqlari tuyoqqa o'xshaydi, uchinchi tirnoqlari yon tomonlarida qo'pol gardish bilan - bu xususiyat boshqa iguanodontlar bilan ham bo'lishgan.[3]

Preacetabular (ya'ni oldida joylashgan kestirib uyasi ) jarayoni ilium gorizontal "yuklash" ga ega, bu jarayonning pastki qismi bilan tekis burchak hosil qiladi. Bundan tashqari, suyakning yuqori qirrasi tashqariga chiqib, iliyum ustida joylashgan chekka hosil qiladi.iskiyum suyakning pastki chetidagi artikulyatsiya. Ushbu xususiyatlar ham ko'rinadi Probaktrosaurus va boshqa iguanodontlar.[3] Kestirib, rozetkaning o'zi iguanodont uchun iliumga nisbatan ancha katta.[4] Iliyumning yuqori yuzasi bo'ylab o'ziga xos gardish mavjud bo'lib, bu belgi noma'lum hadrosauroid bilan birgalikda Woodbine shakllanishi. Orqa tarafdagi iliyum paychalarining postatsetabulyar (ya'ni kestirib, rozetkaning orqasida joylashgan) jarayoni, uning yuqori chetida tanaffuslarsiz. Ustida pubis, oldinga yo'naltirish jarayoni oldinga qarab kengayadi Probaktrosaurus, Gongpoquansaurusva boshqa iguanodontlar. Ishchiumning o'qi kattalarda xarakterli ravishda to'g'ri keladi (balog'at yoshiga etmagan bolalar eguganodontlar orasida ko'proq uchraydigan kavisli o'qga ega) va o'qning pastki qismida oldinga kengaytirilgan siqilgan "botinka" mavjud.[3][7]
Humus kabi suyak suyagi egilgan, lekin tashqariga qarab. Biroq, xarakterli ravishda, femurning pastki yarmi to'g'ri. The to'rtinchi trokanter, femurning yarmida joylashgan, vertikal ravishda baland va uchburchak, o'xshash Probaktrosaurus, Gongpoquansaurus, va boshqa iguanodontlar, ammo ko'proq bazalga o'xshamaydi ornitopodlar. Ulna va radius singari tibia va fibula to'g'ri o'qlarga ega va ular bir-biri bilan aniq ifodalanadi; fibulaning pastki qismidagi qo'pol proektsiya lampochkaga o'xshash ikkita o'rtasida joylashgan kondiller orqasida joylashgan knemial tepalik tibia pastki uchida. Tibianing yuqori qismi qo'pol bo'lib, bu qopqoqning mavjudligini ko'rsatadi xaftaga femur va tibia o'rtasida. Oyoq to'rtta raqamdan iborat. Metakarpalar singari, ikkinchisi metatarsal kavisli, uchinchisi to'g'ri, to'rtinchisi kuchli egri. The falanjlar oyoq juda qattiq, har bir raqamning oxiriga yaqin terminal falanjlari ayniqsa qattiq. Qo'lda ikkinchi va uchinchi tirnoqlarda bo'lgani kabi, oyoqning ikkinchi, uchinchi va to'rtinchi tirnoqlari ham bir tekis keng, tekis va tuyoqqa o'xshaydi.[3]
Tasnifi
Dastlab hadrosaurid identifikatsiyasi

Kirklandning dastlabki tavsifida Eolambiya, u buni a'zosi deb bilgan Hadrosauridae tomonidan belgilanadigan Devid B. Vayshampel, Devid B. Norman, va 1993 yilda Dan Grigoresku. Vayshampel va uning hamkasblari Hadrosauridae-ni aniqlash uchun ettita birlashtiruvchi xususiyatdan foydalanganlar: maxilla ko'tarilish jarayonining yuqoriga kengayishi; kvadratni ajratib turadigan parakuadrat teshiklarining yo'qligi kvadratojugal; joylashuvi burchakli pastki jag'ning ichki yuzasida; surangular teshikchaning yo'qligi; maksillerning tor tishlari; har bir tish tish holatida uch yoki undan ortiq tish borligi; va skapular pichoqning yuqori chetini kamaytirish.[15] Ushbu xususiyatlarning birinchi, beshinchi, oltinchi va ettinchilari tan olingan Eolambiya, qolganlari noma'lum bo'lganligi sababli noma'lum. Kirkland tayinlandi Eolambiya Evshadrosauriyaga, Vayshampel va uning hamkasblari tomonidan umumiy ajdodni kiritish uchun belgilangan Hadrosaurinae (hozirda Saurolophinae) va Lambeosaurinae - hadrosauridlarning ikkita asosiy tarmog'i - va uning barcha avlodlari.[15] Ushbu topshiriq beshta xususiyatga asoslangan edi Eolambiya: premaxilla ustida dentikullar mavjudligi; to'rtburchakning pastki jag 'bilan tor bo'g'imi; tor tish tishlari; har bir tish tishida bitta chekka yoki karina borligi; va humerusda joylashgan deltopektoral tepalik.[2]

Hadrosauridae ichida Kirkland bundan keyin ham ko'rib chiqdi Eolambiya yoki Lambeosaurinae ning bazal a'zosi bo'lish yoki opa-singillar guruhi Lambeosaurinae. U Lambeosaurinae bilan birgalikda turli xil tadqiqotlar davomida aniqlangan beshta xususiyatni aniqladi:[15][16][17][18] premaksilada teshikning yo'qligi; burun teshigining (hech bo'lmaganda qisman) premaxilla tomonidan yopilishi; maxilla ustidagi rafni ishlab chiqish; dumli umurtqalarning juda baland nerv tizmalari; humerusning mustahkamligi; va kattalardagi iskiyaning katta "botinkasi". Kirkland ikkita qo'shimcha xususiyat gavharsizlarni ajratib turishini aniqladi Eolambiya tepalikning rivojlanishi bilan bog'liq bo'lgan boshqa tepalikli lambeozaurinlardan: burun bo'shlig'i ko'z uyasi ustida va parietalning qisqarishi. Agar Eolambiya lambeosaurin deb tan olinishi kerak edi, keyin Lambeosaurinae bu ikkita xususiyatni istisno qilish uchun qayta belgilanishi kerak edi. Shu bilan bir qatorda, agar Eolambiya o'rniga Lambeosaurinae ning singil guruhi bo'lgan, keyin u hadrosauridlar va lambeosaurinlar o'rtasidagi farqga yaqin bo'lgan morozologiyani anglatadi. Ushbu imkoniyat sakrumning pastki qismida yiv borligi bilan qo'llab-quvvatlanadi Eolambiya, bu Kirkland tomonidan hadrosaurinlarning o'ziga xos xususiyati sifatida tan olingan[17][18] - bitta bo'lsa ham yaqinlashuvchi mavjud Ankilozauriya va Ceratopsiya.[2]
Lambeosaurin yaqinligini qo'llab-quvvatlash uchun Eolambiya, Kirkland a filogenetik tahlil; The daraxt tegishli ravishda joylashtirilgan uning tahlili bilan tiklandi Eolambiya eng bazal lambeozaurin sifatida quyida ko'paytiriladi. Ushbu pozitsiyani qo'llab-quvvatlash uchun u keltirgan dalillarga qaramay, Kirkland ham bir-biriga zid ko'rinadigan xususiyatni qayd etdi. U dastlab tasvirlagan balog'atga etmagan ilyumda kuzatilganidek, antitrokanter - iliyumdagi proektsiyani cheklaydi katta trokanter femur - bazal hadrosaurid uchun kutilganidan ancha yomon rivojlangan. Maykl Bret-Surmanning ta'kidlashicha, kattalarda antitrokanter yaxshi rivojlangan bo'lishi mumkin. Boshqa tomondan, olingan lambozaurin Gipakrozavr hatto tuxumdan chiqish bosqichida ham yaxshi rivojlangan antitrokanterga ega. Kirkland bazal hadrosauridlarning antitrokanterni keyinchalik hayotda paydo bo'lgan hadrosauridlarga qaraganda rivojlanishini ta'kidladi. Gipakrozavr.[19] Shunday qilib, u o'sha kattalarni ta'kidladi Eolambiya namunalar bu muammoni hal qiladi.[2]


| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Hadrosaurid bo'lmaganligini aniqlash

2001 yilda Jeyson Xed filogenetik holatini qayta ko'rib chiqdi Eolambiya OMNHda saqlanadigan namunalar asosida. U Kirklandning taxmin qilingan lambeozaurinni ajratib turadigan xususiyatlarini aniqlaganligini tan oldi Eolambiya noto'g'ri yoki shubhali sifatida. Xed tomonidan ko'rsatilgandek, aslida premaksilada foramen mavjud; premaxilla faqat pastki chetdagi burun teshigi bilan chegaralanadi; maksilaning old qismida javondan tashqari ikkita barmoqqa o'xshash jarayonlar mavjud; dumaloq umurtqalarning yuqori nerv umurtqalari lambeozaurinlarda ham uchraydi Ouranosaurus va Bactrosaurus; va iskiyum uchining kengayishi ham sodir bo'ladi Camptosaurus, Iguanodon, Ouranosaurus, Gilmoreosaurus va Bactrosaurus, kengayish ekanligini bildiradi plesiomorfik iguanodonts uchun va ikkinchidan hadrosaurinlarda yo'qolgan.[18] M.L. 1999 yilda Kazanovalar va uning hamkasblari lambozaurinlarni "klubga o'xshash" holatdan farqli o'laroq, iskiyaning uchida "kengaygan oyoq" borligi bilan tavsifladilar,[20] Bosh buni ta'kidladi Korthosaurus va Lambeosaurus[21] iskial maslahatlarga qaraganda ko'proq kengaytirilmagan Mantellisaurus yoki Ouranosaurus.[4][22]

Bosh shuningdek, tishlarning ekanligini ko'rsatdi Eolambiya Kirkland taxmin qilganidan ko'ra kengroq va assimetrik edi; u shuningdek, hadrosauridlardan farqli o'laroq, pastki tishlarda bitta tish holatida faqat ikkita tish borligini ta'kidladi.[4] aslida kattalar namunalari dastlab Kirkland tomonidan ko'rsatilgandek uchta yoki undan ortiq tish holatiga ega.[3] Bosh buni yana ko'rsatdi Eolambiya sakkizta va undan ko'prog'idan hosil bo'lgan hadrosauridlardan farqli o'laroq, ettita sakral o'murtqa ega edi va uning antitrokanteri, aslida, juda kam rivojlangan edi. Ouranosaurus va Altirhinus.[23] OMNH namunalarini o'rganish shuni ko'rsatdiki, iliumning postatsetabulyar mintaqasi alohida jarayonni hosil qilmagan va femurning pastki qismidagi artikulyatsiyalar unchalik ko'zga tashlanmaydi, ikkalasi ham ajralib turadi Eolambiya hadrosauridlar kabi ko'proq olingan iguanodontlardan. Shunday qilib, Bosh shunday xulosaga keldi Eolambiya edi a bazal Hadrosauridadan tashqarida bo'lgan Hadrosauroidea a'zosi, filogenetik tahlil bilan tasdiqlangan xulosaga ko'ra uni hadrosaurid bo'lmagan hadrosauroid bilan chambarchas bog'liq Probaktrosaurus.[4]
Keyingi tadqiqotlar

2004 yilda Norman qo'shildi Eolambiya birinchi marta iguanodontsni filogenetik tahlilida, ikkinchi nashrida iguanodontsga bag'ishlangan bobning bir qismi sifatida Dinozavrlar. U buni topdi Eolambiya singil guruh edi Altirhinus, ikkalasi ham bazalroq bo'lganligi bilan Protohadros, Probaktrosaurusyoki Hadrosauridae.[24] Xuddi shu jildda Horner, Vayshampel va Ketrin Forster hadrosauridlarning alohida filogenetik tahlilini o'tkazdi, shu jumladan Eolambiya. Ular topdilar Eolambiya singari guruh bo'lsa ham, yana adrosaurid bo'lmagan hadrosauroid sifatida Protohadros. Garchi Head Hadrosauridae-ni umumiy xususiyatlarga asoslanib, o'z ichiga olgan holda qayta aniqlagan bo'lsa-da Protohadros,[4][25] Xorner va uning hamkasblari taksonga asoslangan ta'rifni chiqarib tashladilar Protohadros va shunday qilib Eolambiya. Shuningdek, ular differentsiyalashgan qo'shimcha xususiyatlarni aniqladilar Eolambiya hadrosauridlardan: tish tishlarida qo'pol dentikulalar mavjud va koronoid jarayon zaif kengaygan.[26]

Qayta tiklangan filogenetik pozitsiyalardagi farq Eolambiya keyingi yillarda davom etdi. 2009 yil tavsifida Levnesoviya, Xans-Diter Sues va Aleksandr Averianov buni topdi Protohadros ga nisbatan oraliq pozitsiyani egallagan Altirhinus va Probaktrosaurus, singil guruh bo'lib Fukuisaurus.[27] Ayni paytda Horner va uning hamkasblari natijalariga ko'ra Albert Prieto-Markes va Mark Norell 2010 yilda topilgan Eolambiya o'rniga uning singil guruhi bo'lgan Protohadros. Shuningdek, u xususiyatlarini qayd etdi Eolambiya hadrosauridlarga yaqinlashadigan: bitta tish karinasi borligi; oldingi kvadratning deyarli kvadratik tashqi burchagi; va kvadratojugal chuqurchaning (to'rtburchak bilan birikadigan) o'rtasi to'rtburchak yuzasining taxminan yarmida joylashgan (yuqoridan suyak balandligining 60% dan kamrog'ida joylashgan).[28] 2012 yilda Xolts tasniflangan Eolambiya ning ibtidoiy a'zosi sifatida Styracosterna bilan birga Altirhinus.[14]
2012 yilda McDonald iguanodontlarning filogenetik tahlilini o'tkazdi, ular yangi ma'lumotlarni o'z ichiga olgan Eolambiya namunalar. Oldingi mualliflardan farqli o'laroq, u buni topdi Eolambiya ning opa-singil guruhi edi Probaktrosaurus. Ushbu yaqin munosabatlar ikkita umumiy xususiyatga asoslangan edi: to'rtburchak yuqoriga qarab, orqaga qarab egilgan; skapular pichoqning yuqori va pastki chetlari deyarli parallel.[3] Wenjie Zheng va uning hamkasblari qo'shildi Jintasaurus 2014 yilda guruhga,[29] Xose Gaska va uning hamkasblari esa dastlabki guruhni 2015 yilda tiklagan.[30] Shu bilan birga, McDonald FMNH namunalarining 2017 yilgi tavsifiga tayyorgarlik ko'rish jarayonida o'zining filogenetik ma'lumotlar to'plamini keng ko'lamda qayta ko'rib chiqdi. Horner va uning hamkasblari, shuningdek Prieto-Markes va Norell singari, McDonald va uning hamkasblari buni aniqladilar Eolambiya ning opa-singil guruhi edi Protohadros, ikkalasi ham bazal sifatida joylashtirilgan hadrosauromorflar (natijalar quyida keltirilgan).[7] McDonald ma'lumotlar to'plamining 2012 yilgi versiyasidan foydalangan holda, Frantsisko Verdu va uning hamkasblari mustaqil ravishda shu kabi natijani tikladilar,[31][32] bilan Eolambiya bilan chambarchas bog'liq Protohadros va Jeyavati; ammo, McDonald va uning hamkasblari ikkinchisini 2017 yilda ko'proq olingan deb topdilar.[7]

| Hadrosauriformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2018 yilda Terri Geyts va uning hamkasblari yangi iguanodotian turini tasvirlab berishdi Choyrodon, xuddi shu joydan Altirhinus, o'tmishda qarindoshi deb hisoblangan Eolambiya. Norman (2015) matritsasi asosida ularning filogenetik tahlillari topildi Choyrodon ning singlisi taksoni bo'lish Eolambiya. Eolambiya'qarindoshlari odatda tiklanadi Protohadros ushbu matritsada bo'lmagan. Ushbu singil munosabatlarni uchta xususiyat qo'llab-quvvatladi: trapetsiya shaklida oksiput va tishlarning ikkita umumiy xususiyati. Ushbu nasllarning boshqa hadrosauriformesga nisbatan joylashishi antorbital fenestraning mavjud yoki yo'q deb kodlanganligiga bog'liq. Choyrodon; voyaga etmaganlar uchun namunada namunalar mavjud, ammo kattalar yopiq bo'lishi ehtimolini inkor etib bo'lmaydi. Quyidagi ikkita daraxt (matnda Geyts va uning hamkasblari tomonidan belgilangan xochlar bilan belgilangan) natijalarni o'z navbatida mavjud va yo'q deb kodlangan holda ko'rsatadi, birinchisi katta polotomiya:[33][tekshirib bo'lmadi ]
Topologiya A: Antorbital fenestra mavjud bo'lgan joyda kodlangan Choyrodon[33]
| B topologiyasi: Antorbital fenestra tashqi tomondan yopiq deb kodlangan Choyrodon[33]
|
Paleobiologiya
Ehtimol, tegishli bo'lgan tuxum qobig'i Eolambiya izolyatsiya qilingan tishlar bilan bir qatorda turli joylarda topilgan. Ular retikulyar (to'rga o'xshash) sirt naqshlariga ega va qalinligi 2 millimetrga teng (0,079 dyuym).[2]
O'sish

Voyaga etmaganlar va kattalar uchun turli xil namunalardan ma'lum bo'lish, o'zgarishlar Eolambiya o'sib ulg'ayganligi sababli hujjatlashtirilgan. Maksilladagi tishlarning soni yoshga qarab o'sib bordi, balog'atga etmagan bolada 23 tadan kattalarda 33 taga etdi. Shunga o'xshash o'sish stomatologik tishlar, o'spirinlarda 18-22 yoshdan kattalarda 25-30 gacha. Voyaga etmaganlarga tish tishlarining tojlarida ikkilamchi tizmalar etishmasa, zaif ikkilamchi tizmalar kattalarda uchraydi.[3] Ishkiumning "botinkasi" ning kattaligi ham Kirkland tomonidan tasdiqlangan yoshga qarab oshdi.[2] FMNH tomonidan kashf etilgan namunalar bilan hujjatlashtirilgandek, iskiyya milining tekisligida o'zgarishlar ham bo'lishi mumkin.[7]

A natijalari gistologik FMNH PR 3847 namunasidagi qovurg'ada o'tkazilgan tahlillar McDonald va uning hamkasblari tomonidan 2017 yilda xabar qilingan. suyaklarni qayta qurish qovurg'aning ichki matritsasida paydo bo'lgan. The medullyar bo'shliq, qaysi joylashgan ilik, ko'plab bo'shliqlar mavjudligi bilan belgilanadi yoki lakuna. Suyakning tashqi yuzasida, osteonlar suyakning ichki qismini deyarli butunlay o'zgartirgan; suyakning qatlamli o'sishi suyakning ichki qismida ko'proq ko'rinadi, bu erda beshta hibsga olingan o'sish yo'nalishlari (LAG) ko'rinadi. Oltinchi LAG medullar mintaqasida bo'lishi mumkin, ammo nisbatan cheklangan, bu esa bu identifikatsiyani shubha ostiga qo'yadi.[7]
LAGlarni suyaklarni qayta qurish yo'li bilan yo'q qilish mumkinligi sababli, bu aslida rivojlangan LAGlarning haqiqiy sonini kam baholashni anglatadi; Makdonald va uning hamkasblari ko'rinadigan LAGlar orasidagi masofani o'rganib, uch yoki to'rtta etishmayotgan LAGni taxmin qilishdi. Bu hayvon o'lganda sakkiz-to'qqiz yoshda bo'lganligini ko'rsatadi. LAGlar orasidagi masofa tashqi yuzaga qarab sezilarli darajada pasayadi, bu o'sish sekinlashayotganini ko'rsatadi, ammo tashqi fundamental tizimning etishmasligi (bir-biridan zich joylashgan LAGlar) o'sish hali to'xtamaganligini ko'rsatadi. Bu bachadon bo'yi va dumg'aza umurtqasidagi sentra va asab orqa miya orasidagi tikilmagan tikuvlarga va sakral tsentra va ularga mos keladigan qovurg'alar orasidagi tikilmagan tikuvlarga mos keladi.[34] Umuman olganda, o'sish sur'ati Eolambiya bilan solishtirish mumkin bo'lgan ko'rinadi Gipakrozavr va Mayasaura yetti yoki sakkiz yildan so'ng etuklikka erishdi.[7][35][36]
Paleoekologiya
Atrof muhit va taponomiya
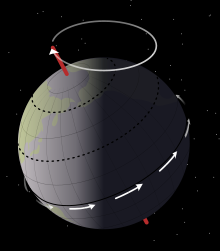
Sedimentologik Mussentuchit a'zosi ma'lumotlari shuni ko'rsatadiki Eolambiya yomon quriganlikda yashagan toshqin suv toshqini, katta, ko'p yillik ko'llar tizimi atrofida. Ko'llar suv sathining yuqori va past darajalarida o'zgarib turardi Milankovichning tsikllari taxminan 21000 yil, Yer tomonidan yuzaga kelgan eksenel prekretsiya. Ushbu tsikllar hozirgi tog 'jinslari konlarida loyli qumtosh va loy toshlarining o'zgaruvchan qatlamlari bilan aks etadi. During dry periods, the exposed lakebed formed a broad beach devoid of plants. The cracked mud of the lakebed produced loy parchalari, while organic debris and scavenged carcasses were scattered around the water's edge as the lake progressively retreated. Meanwhile, during periods in which the sea level rose, small rivers invaded the floodplain, which qayta ishlangan and displaced the debris that had been deposited during the dry season.[6]

Most of the fossils discovered in the Mussentuchit are scattered and disarticulated, with intact skeletons being relatively rare. Although the lakes did not actively destroy bones, it was not particularly conducive to good preservation either due to the long duration of the burial process. Before burial, scavengers could have disrupted the skeletal material, which is evident in the severed Eolambiya tibiae bearing tooth marks discovered in the Cifelli #2 quarry. The regression of the lake could have occasionally created bog ', but they would not have been permanent enough to entrap and preserve larger vertebrates. Action by water currents seems to have played a role in the deposition of Eolambiya bones in the Cifelli #2 quarry, where the bones are largely deposited at 28°, 69°, 93°, 131° and 161° azimut (i.e. relative to north). These directions would have been parallel or sub-parallel to the edges of the lake, indicating orientation by lake currents, except for the 69° and 93° azimuths, which probably represent river currents.[6]
The abundant plant debris, combined with the absence of cho'kish (playa) deposits, indicates that the local climate was relatively humid during the deposition of the Mussentuchit. This is in marked contrast to earlier deposits in the area, which are ohakli (chalky), indicating periods of aridity lasting up to 10,000 years. Paleoklimatik reconstructions support these interpretations, with the encroachment of the Mowry Sea from the north (a process which would eventually form the G'arbiy ichki dengiz yo'li ) resulting in arid (Mid-latitude Continental Interior) climates being replaced by more humid (Mid-latitudeEast Coast) climates.[37] With a paleolatitude of 46° N, the Mussentuchit would have had a warm mean surface temperature of 40–45 °C (104–113 °F), about 6–8 °C (43–46 °F) warmer than the present day, according to umumiy aylanish modellari.[6][38]
Ekotizim

Crocodylomorphs are abundant in the Mussentuchit deposits, being represented largely by teeth. The largest known teeth have been referred to the Pholidosauridae;[a] they are broad with rounded bases, and some of them bear longitudinal ridges.[7] These large crocodilians would have been capable of preying upon at least juvenile Eolambiya suv bo'yida The abundance of juvenile Eolambiya specimens in the lakeside fossil material supports this hypothesis; kattaroq Eolambiya would have been invulnerable to crocodilians.[6] Smaller crocodilian teeth are also present, with two distinct morphotypes: an ovoid form bearing ridges, which has been referred to two different species of Bernissartiya;[6][a] and a compressed, triangular form bearing heavy ornamentation, which has been attributed to the Atoposauridae.[12][40][a] However, given the uniform small size of these teeth, and the co-occurrence of these morphotypes in some crocodilians,[39] they may belong to the same species.[7]

Kattaroq Eolambiya would have been vulnerable[6] to the large neovenatorid teropod Siats,[41] which was discovered by expeditions from the FMNH.[7] A variety of smaller maniraptoran theropods are also present in the Mussentuchit, which would have functioned as scavengers; their remains include teeth attributed to the Dromaeosauridae, Paronixodon va Richardoestesia.[40] As for herbivorous dinosaurs, the nodosaurid Animantarx was discovered in Carol's Site, near the type specimen of Eolambiya.[42] Teeth belonging to more basal ornithopods have also been discovered, with similar teeth having been referred to the Pachycephalosauria or to toothed birds by Cifelli in 1999.[6][40] More complete remains referred to two different ornithopods – an orodromin and a basal iguanodont – remain unpublished.[43] Gigant oviraptorosaur has also been discovered,[44] alongside teeth from ceratopsians and sauropodlar.[7][40]
Fish, which may have been preyed upon by smaller crocodilians, are represented by teeth and scales. Ular tarkibiga a myliobatiform ray referred to the genus Baibisha;[40] an indeterminate member of the Neopterygii; a semionotid referred to the genus Lepidotlar; a gar bog'liq bo'lgan Atractosteus;[7] a pycnodontid referred to the genus Stephanodus; va noaniq bowfin. Belgilanmagan kaudatanlar (salamanders) and mammals are also present. As for plants, pollen samples indicate a tiered forest surrounding the lakes of the Mussentuchit, containing a soyabon ning daraxt ferns, araucarias va ignabargli daraxtlar, accompanying an undergrowth of ferns, kichik gimnospermlar va gullarni o'simliklar. Algae would also have been present in the waters of the lakes.[9][45] These plants would have provided an abundant source of food for Eolambiya.[6]
Paleobiogeografiya

Kirkland, Cifelli, and colleagues noted that the fauna of the Mussentuchit – iguanodonts, pachycephalosaurs, and ceratopsians – bears strong similarities to contemporary Asian faunas. They proposed that Eolambiya was part of an influx of Asian dinosaurs into North America during the Cenomanian, which supplanted the earlier low-diversity native fauna.[2][12] This hypothesis is supported by the close relationship between Eolambiya va ham Probaktrosaurus yoki Fukuisaurus, which have respectively been recovered by the phylogenies of Head and Sues & Averianov.[3][4][27]
However, the results of Horner and colleagues, Prieto-Márquez, and McDonald and colleagues, which consider the North American Protohadros ning eng yaqin qarindoshi bo'lish Eolambiya, contradict this hypothesis.[7][26][28][46] This alternative interpretation suggests that the common ancestor of Eolambiya, Protohadros, and more derived hadrosauroids was not exclusively Asian, but instead distributed across Asia and North America.[3] McDonald and colleagues further suggested that Eolambiya va Protohadros represent a localized group of hadrosauroids endemik to North America, much like other groups of styracosternans throughout the Cretaceous. Such contemporary groups included a central Asian group of Shuangmiaosaurus, Zhanghenglong va Plesiohadros.[7]
Adabiyotlar
- ^ Galton, Piter; Jensen, James A. (1979). "Galton, P. M., & Jensen, J. A. (1979). Remains of ornithopod dinosaurs from the Lower Cretaceous of North America" (PDF). Brigham Young universiteti geologiya tadqiqotlari. 25 (3): 1–10. ISSN 0068-1016.
- ^ a b v d e f g h men j k l m Kirkland, J.I. (1998). "A new hadrosaurid from the upper Cedar Mountain Formation (Albian-Cenomanian: Cretaceous) of eastern Utah – the oldest known hadrosaurid (lambeosaurine?)". Lukasda S.G .; Kirkland, J.I .; Estep, J.W. (tahr.). Lower and Middle Cretaceous Terrestrial Ecosystems. Nyu-Meksiko Tabiat Tarixi va Ilmiy Muzeyi. 14. pp. 283–295.
- ^ a b v d e f g h men j k l m n o p q r s t McDonald, AT; Qush, J .; Kirkland, J.I .; Dodson, P. (2012). "Osteology of the Basal Hadrosauroid Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation of Utah". PLOS ONE. 7 (10): e45712. Bibcode:2012PLoSO...745712M. doi:10.1371/journal.pone.0045712. PMC 3471925. PMID 23077495.
- ^ a b v d e f g Boshliq, J.J. (2001). "A reanalysis of the phylogenetic position of Eolambia caroljonesa (Dinosauria, Iguanodontia)". Umurtqali hayvonlar paleontologiyasi jurnali. 21 (2): 392–396. doi:10.1671/0272-4634(2001)021[0392:AROTPP]2.0.CO;2.
- ^ Kirkland, J.I .; Burge, D. (1994). A large primitive hadrosaur from the Lower Cretaceous of Utah. Fifty-Fourth Annual Meeting, Society of Vertebrate Paleontology. Umurtqali hayvonlar paleontologiyasi jurnali. 14 (3). p. 32A. JSTOR i406457.
- ^ a b v d e f g h men j k l m Garrison, J.R. Jr.; Brinkman, D.; Nichols, D.J .; Layer, P.; Burge, D.; Thayn, D. (2007). "A multidisciplinary study of the Lower Cretaceous Cedar Mountain Formation, Mussentuchit Wash, Utah: a determination of the paleoenvironment and paleoecology of the Eolambia caroljonesa dinosaur quarry". Bo'r davridagi tadqiqotlar. 28 (3): 461–494. doi:10.1016/j.cretres.2006.07.007.
- ^ a b v d e f g h men j k l m n o p q r s McDonald, AT; Geyts, T.A .; Zanno, L.E .; Makovicky, P.J. (2017). "Anatomy, taphonomy, and phylogenetic implications of a new specimen of Eolambia caroljonesa (Dinosauria: Ornithopoda) from the Cedar Mountain Formation, Utah, USA". PLOS ONE. 12 (5): e0176896. Bibcode:2017PLoSO..1276896M. doi:10.1371/journal.pone.0176896. PMC 5425030. PMID 28489871.
- ^ Kreyg, LC (1981). "Lower Cretaceous rocks, southwestern Colorado and southeastern Utah". Rocky Mountain Association of Geologists 1981 Field Conference: 195–200.
- ^ a b Tschudy, R.H.; Tschudy, B.D.; Kreyg, LC (1984). Palynological evaluation of Cedar Mountain and Burro Canyon formations, Colorado Plateau (Hisobot). US Geological Survey, Professional Paper. 1821. AQSh Geologik xizmati. 1-24 betlar.
- ^ Nichols, D.J .; Sweet, A.R. (1993). "Biostratigraphy of Upper Cretaceous nonmarine palynofloras in a north-south transect of the Western Interior Basin". In Caldwell, W.G.E.; Kauffman, E.G. (tahr.). Evolution of the Western Interior Basin. Geological Association of Canada, Special Paper. 39. Kanada geologik assotsiatsiyasi. pp. 539–584.
- ^ Yosh, R.G. (1960). "Dakota Group of Colorado Plateau". Amerika neft geologlari byulleteni assotsiatsiyasi. 44 (2): 156–194. doi:10.1306/0bda5fd2-16bd-11d7-8645000102c1865d.
- ^ a b v Cifelli, R.L.; Kirkland, J.I .; Weil, A.; Deino, A.L.; Kowallis, B.J. (1997). "High-precision 40Ar/39Ar geochronology and the advent of North America's Late Cretaceous terrestrial fauna". Milliy fanlar akademiyasi materiallari. 94 (21): 11163–11167. Bibcode:1997PNAS...9411163C. doi:10.1073/pnas.94.21.11163. PMC 23402. PMID 9326579.
- ^ Pol, GS (2010). "Ornithischians". Dinozavrlar uchun Princeton Field Guide. Prinston: Prinston universiteti matbuoti. p. 327. ISBN 9780691167664.
- ^ a b Xolts, T.R. (2012). "Genus List for Holtz (2007) Dinozavrlar" (PDF). Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages by Thomas R. Holtz, Jr., illustrations by Luis Rey. Department of Geology, University of Maryland. Olingan 30 avgust, 2017.
- ^ a b v Vayshampel, D.B.; Norman, DB .; Grigorescu, D. (1993). "Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur". Paleontologiya. 36 (2): 361–385.
- ^ Sereno, P.C. (1986). "Qushlarga tashlangan dinozavrlarning filogeniyasi (Ornithischia ordeni)". Milliy geografik tadqiqotlar. 2: 234–256.
- ^ a b Brett-Surman, M.K. (1989). Hadrosauridalarni qayta ko'rib chiqish (Reptiliya: Ornithischia) va ularning Campanian va Maastrichtian davrida rivojlanishi (PDF) (Fan nomzodi). Vashington D.C .: Jorj Vashington universiteti. pp. 1–373.
- ^ a b v Vayshampel, D.B.; Horner, J.R. (1990). "Hadrosauridae". Vayshampelda, D.B.; Dodson, P.; Osmolska, H. (tahrir). Dinozavrlar (1-nashr). Berkli: Kaliforniya universiteti matbuoti. 534-561 betlar.
- ^ Xorner, JR .; Currie, P.J. (1994). "Embryonic and neonatal morphology and ontogeny of a new species of Gipakrozavr (Ornithischia, Lambeosauridae) from Montana and Alberta". Duradgorda K.; Hirsch, K.F.; Horner, J.R. (eds.). Dinozavrlarning tuxumlari va chaqaloqlari. Kembrij: Kembrij universiteti matbuoti. pp. 312–336. ISBN 9780521567237.
- ^ Casanovas, M.L.; Pereda Suberbiola, X.; Santafe, J.V.; Weishampel, D.B. (1999). "First lambeosaurine hadrosaurid from Europe: palaeobiogeographical implications". Geologik jurnal. 136 (2): 205–211. Bibcode:1999GeoM..136..205C. doi:10.1017/s0016756899002319.
- ^ Lull, R.S.; Wright, N.E. (1942). "Hadrosaurian Dinosaurs of North America". Amerika Geologik Jamiyati Maxsus Hujjatlar. 40: 1–242. doi:10.1130/SPE40-p1.
- ^ Norman, D.B. (1986). "On the anatomy of Iguanodon atherfieldensis (Ornithischia: Ornithopoda)". Bulletin de l'Institut Royal des Sciences Naturelles de Belgique. 56: 281–372.
- ^ Norman, D.B. (1998). "On Asian ornithopods (Dinosauria: Ornithischia). 3. A new species of iguanodontid dinosaur". Linnean Jamiyatining Zoologik jurnali. 122 (1): 291–348. doi:10.1111/j.1096-3642.1998.tb02533.x.
- ^ Norman, D.B. (2004). "Basal Iguanodontia". Vayshampelda, D.B.; Dodson, P.; Osmolska, H. (tahr.) Dinozavrlar (2-nashr). Berkli: Kaliforniya universiteti matbuoti. 413-437 betlar.
- ^ Boshliq, J.J. (1998). "A new species of basal hadrosaurid (Dinosauria, Ornithischia) from the Cenomanian of Texas". Umurtqali hayvonlar paleontologiyasi jurnali. 18 (4): 718–738. doi:10.1080/02724634.1998.10011101.
- ^ a b Xorner, JR .; Vayshampel, D.B.; Forster, C.A. (2004). "Basal Iguanodontia". Vayshampelda, D.B.; Dodson, P.; Osmolska, H. (tahr.) Dinozavrlar (2-nashr). Berkli: Kaliforniya universiteti matbuoti. pp. 438–463.
- ^ a b Sues, H.-D .; Averianov, A. (2009). "O'tgan bo'r davridan yangi bazal hadrosauroid dinozavr va o'rdak parchalanadigan dinozavrlarning erta nurlanishi". Qirollik jamiyati materiallari B. 276 (1667): 2549–2555. doi:10.1098 / rspb.2009.0229. PMC 2686654. PMID 19386651.
- ^ a b Prieto-Markes, A.; Norell, MA (2010). "Anatomiya va aloqalari Gilmoreosaurus mongoliensis (Dinosauria: Hadrosauroidea) from the Late Cretaceous of Central Asia". Amerika muzeyi Novitates. 3694: 1–49. doi:10.1206/3694.2. S2CID 56372891.
- ^ Zheng, W.; Jin X.; Shibata M.; Azuma, Y. (2014). "An early juvenile specimen of Bolong yixianensis (Ornithopoda: Iguanodontia) from the Lower Cretaceous of Ningcheng County, Nei Mongol, China". Tarixiy biologiya. 26 (2): 236–251. doi:10.1080/08912963.2013.809347. S2CID 129081459.
- ^ Gasca, J.M.; Moreno-Azanza, M.; Ruiz-Omeñaca, J.I.; Kanudo, J.I. (2015). "New material and phylogenetic position of the basal iguanodont dinosaur Delapparentia turolensis from the Barremian (Early Cretaceous) of Spain". Iberian Geology jurnali. 41 (1): 57–70. doi:10.5209/rev_jige.2015.v41.n1.48655.
- ^ Verdú, F.J.; Royo-Torres, R .; Kobos, A .; Alcalá, L. (2015). "Perinates of a new species of Iguanodon (Ornithischia: Ornithopoda) from the lower Barremian of Galve (Teruel, Spain)". Bo'r davridagi tadqiqotlar. 56: 250–264. doi:10.1016/j.cretres.2015.05.010.
- ^ Verdú, F.J.; Royo-Torres, R .; Kobos, A .; Alkala, L. (2017). "New systematic and phylogenetic data about the early Barremian Iguanodon galvensis (Ornithopoda: Iguanodontoidea) from Spain". Tarixiy biologiya. 30 (4): 437–474. doi:10.1080/08912963.2017.1287179. S2CID 89715643.
- ^ a b v Geyts, T.A .; Tsogtbaatar, K .; Zano, L.E.; Chinzorig, T .; Watabe, M. (2018). "A new iguanodontian (Dinosauria: Ornithopoda) from the Early Cretaceous of Mongolia". PeerJ. 6: e5300. doi:10.7717/peerj.5300. PMC 6078070. PMID 30083450.
- ^ Brochu, Kaliforniya (1994). "Closure of neurocentral sutures during crocodilian ontogeny: Implications for maturity assessment in fossil archosaurs". Umurtqali hayvonlar paleontologiyasi jurnali. 16 (1): 49–62. doi:10.1080/02724634.1996.10011283.
- ^ Xorner, JR .; de Rikkles, A .; Padian, K. (1999). "Variation in dinosaur skeletochronology indicators: implications for age assessment and physiology". Paleobiologiya. 25 (3): 295–304. doi:10.1017/S0094837300021308.
- ^ Woodward, H.N.; Freedman Fowler, E.A.; Farlow, J.O .; Horner, J.R. (2015). "Mayasaura, a model organism for extinct vertebrate population biology: a large sample statistical assessment of growth dynamics and survivorship". Paleobiologiya. 41 (4): 503–527. doi:10.1017/pab.2015.19. S2CID 85902880.
- ^ Elder, W.P.; Kirkland, J.I. "Cretaceous Paleogeography of the Southern Western Interior Region". In Caputo, M.V.; Peterson, J.A.; Franczyk, K.J. (tahr.). Mesozoic Systems of the Rocky Mountain Region, USA. Denver: Rocky Mountain Section SEPM. pp. 415–440.
- ^ Glancy, T.J. Jr.; Artur, M.A .; Barron, E.J.; Kaufman, E.G. (1993). "A paleoclimate model for the North American Cretaceous (Cenomanian-Turonian) epicontinental sea". In Caldwell, W.G.E.; Kauffman, E.G. (tahr.). Evolution of the Western Interior Basin. Geological Association of Canada, Special Papers. 39. Kanada geologik assotsiatsiyasi. 219-241 betlar.
- ^ a b Irmis, RB .; Hutchison, J.H.; Sertich, JJW; Titus, A.L. (2013). "Crocodyliforms from the Late Cretaceous of Grand Staircase-Escalante National Monument and Vicinity, Southern Utah, USA". Titusda A.L .; Loewen, MA (tahrir). At the Top of the Grand Staircase: the Late Cretaceous of Southern Utah. Bloomington: Indiana universiteti matbuoti. pp. 424–444. ISBN 9780253008961.
- ^ a b v d e Cifelli, R.L.; Nydam, R.L.; Gardner, J.D.; Weil, A.; Eaton, JG .; Kirkland, J.I .; Madsen, S.K. (1999). "Medial Cretaceous vertebrates from the Cedar Mountain Formation, Emery County, Utah: the Mussentuchit local fauna". In Gillette, D. (ed.). Yuta shtatidagi umurtqali hayvonlar paleontologiyasi. Utah Geological Survey Miscellaneous Publications. 99–1. pp. 219–242.
- ^ Zanno, L.E .; Makovicky, PJ (2013). "Neovenatorid theropodlar - Shimoliy Amerikaning so'nggi bo'r davridagi tepalik yirtqichlari". Tabiat aloqalari. 4: 2827. Bibcode:2013 NatCo ... 4.2827Z. doi:10.1038 / ncomms3827. PMID 24264527.
- ^ Duradgor, K .; Kirkland, J.I .; Burge, D.L.; Bird, J. (1999). "Ankylosaurs (Dinosauria: Ornithischia) of the Cedar Mountain Formation, Utah, and their stratigraphic distribution". In Gillette, D. (ed.). Yuta shtatidagi umurtqali hayvonlar paleontologiyasi. Utah Geological Survey Miscellaneous Publications. 99–1. 243–251 betlar.
- ^ Zanno, L.E .; Makovicky, PJ (2016). A New Species of Early Diverging Ornithopod Increases the Paleobiodiversity of Herbivorous Dinosaurs in Late Cretaceous Ecosystems in North America. Society of Vertebrate Paleontology 76th Annual Meeting. Salt Lake City: Society of Vertebrate Paleontology. p. 256.
- ^ Makovicky, PJ .; Zanno, L.E. (2014). Kirkland, J.I .; Hunt-Foster, R.; Liggett, G.A.; Trujillo, K. (eds.). New Discoveries Add to the Diversity of the Mussentuchit Member (Cedar Mountain Formation) Dinosaur Fauna. Mid-Mesozoic: The Age of Dinosaurs in Transition (PDF). Fruita: Utah Geological Survey. 70-71 betlar.
- ^ Ravn, R.L. (1995). "Miospores from the Muddy Sandstone (Upper Albian), Wind River Basin, Wyoming, USA". Paleontografiya Abteilung B. 234 (3): 41–91.
- ^ Prieto-Márquez, A. (2010). "Global historical biogeography of hadrosaurid dinosaurs". Linnean Jamiyatining Zoologik jurnali. 159 (2): 503–525. doi:10.1111/j.1096-3642.2010.00642.x.
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||






